
生命的奇迹和造物主的伟大智慧
第二章 超凡绝伦的生命围墙——细胞膜
作者:孟荪
安得广厦千万间,
大庇天下寒士俱欢颜,
风雨不动安如山。
——杜甫
尽管还拿不出任何像样的证据,表明地球原始状态下能从无机物产生有机生命小分子。但毕竟生命出现了,暂且先把这个问题搁一边,姑且算是做无米之炊吧,以下各章将选择一些例子,说明生命之道的难度是何等之大。
这一章先从最简单的说起。
所有生命体都必须将维持活命的各种有效成分留在体内,不让随意流失,因此,必须有一个围好的“家”。今天的生命科学公认,最原始的生命是单细胞生物。细胞膜就仿佛是这个“家”的墙,给细胞围出了一个“家”,是保证细胞存活的最基本条件。细胞的直径一般只有十多微米,相当于1%毫米,只在显微镜下才能看见。而细胞膜厚度仅仅几个纳米,万分之一毫米左右,普通光学显微镜连半点儿影子都看不到。
所有固体无机物的岩石、矿物、冰块等等都是块状的,成不了膜。我们生活中接触到的薄膜,无论是纸张或是塑料膜等等都是用有机产品生产的。可是,原始生命哪来这些有机产品呢?看来我们又一次面临无米之炊的尴尬境地。不过暂且把这个问题搁一边,先看看细胞膜具有哪些基本特性。
一、细胞膜基本分子需要的双重特性
我们来看一下细胞膜应该满足哪些基本要求:
首先,生命细胞生活在水中,细胞内部也充满水,细胞必须不断从水环境中获得各种营养物质,同时又将各种代谢废物排到细胞外。所以包裹细胞的膜必须内外两面都能与水保持密切接触,也就是说必须内外都能亲近水的。但是,如果细胞膜只是亲水的话,由于细胞内有大量高浓度的生命分子,而细胞外的浓度则低得多。水分是会自动从低浓度渗透到高浓度处。这样大量的水份会就从细胞外渗透到细胞内,很快就把细胞撑破。因此,细胞膜还应该具有拒水的特征,以便能将水份拒之门外。所以,细胞膜对水必须“半推半就”,又推又就的双重性。
另方面,细胞膜必须有一定坚韧性。常温下分子热运动的速度与子弹差不多,所以,细胞几乎始终处于枪林弹雨之中,如果细胞膜不够坚韧的话,很快就会被击穿。如果细胞膜分子彼此之间依靠化学键彼此连接,那就会牢固像钢筋水泥墙。我们生活中牢固的薄膜很常见,如聚乙烯、聚丙烯,或尼龙薄膜,分子之间彼此用化学键连接,十分牢固,连空气都难通过。但是,细胞还必须通过细胞膜上的许许多多通道让生命活动分子能进进出出,因此,细胞膜必须有一定的可“穿凿”性,以便在上面穿凿许许多多通道。如果细胞膜很难“凿”开,无法开辟通道,就将把细胞活活闷死。但是,如果不用化学键相互连接的话,那又怎样保持像防弹背心那样牢固?没用化学键链接的颗粒,例如,一堆灰尘,风一吹就散了。
细胞膜既要亲水又要拒水,既要“坚韧”又要易于“穿凿”,必须同时具有这样截然相反的两种特性,如何能做到呢?
先谈谈亲水和拒水。亲水膜不少见,例如纸张,水一溅上去就渗透进去了,湿漉漉的。拒水的膜也不少,例如物体涂了油膜的表面和各种塑料薄膜等等,水泼上去,成了一滴滴水珠。都只能满足单一要求。
现代科技也发明了不少既拒水又亲水的材料,洗涤剂就是典型。工业生产的洗涤剂的分子基本上都是一个脑袋拖一根尾巴,脑袋一端亲水,尾巴那一端拒水但能亲油污。于是亲油污的尾巴粘住了油污,亲水的脑袋就往水那边钻去,于是就将油性污垢拉入水中。
如果将一小滴洗涤剂滴入水中,洗涤剂分子的亲水脑袋都向外迎着水,尾巴都统统被挤到中央。在周围水的亲和压力下,洗涤剂分子会被紧密挤压在一起,凝聚成直径200纳米左右的实心小球。小球内没有一点儿空间去容纳别的分子。如果将很多洗涤剂投入水中,则会全都平铺到水面上,亲水脑袋朝下贴着水面,拒水尾巴一律朝上迎着空气。所以,通常的洗涤剂分子虽然同时具有拒水和亲水两种特性,却不足以作为细胞膜的材料(图1)。
到这里,建议阁下停下来思考一番,看看您能有何妙招。
恐怕很难吧。
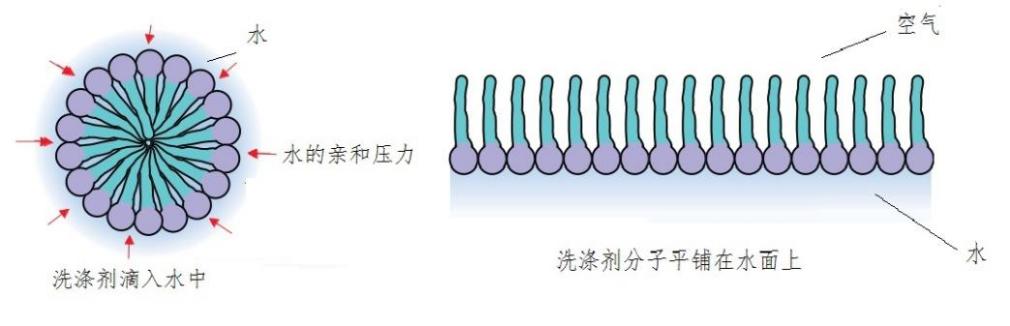
图1 洗涤剂分子不能成为细胞膜材料
现在来看看生命的解决之道。
构建细胞膜的最基本成分是一类叫做“磷脂”的化学分子,占细胞膜重量的20%-80%,相当于砌墙的砖块。它们结构是一个 “脑袋”拖两根长长的尾巴。它们的脑袋喜欢亲近水;而那两根尾巴却相反,是拒水的。洗涤剂分子只有一根尾巴,而磷脂分子有两根,多了一根。别看是仅仅多了一根,却有极巧妙的妙思。因为这两根尾巴叉得比较开,难以将它们挤成不留空隙的实心球,就可以留有内部空间了。瞧,多妙啊!
不过,有可能会产生另外的一个问题,那就是如果水分子钻进内部空间,会迫使磷脂的拒水尾巴尽量躲远远的,这一来,磷脂膜就被解体了。
然而,细胞膜的磷脂却有巧妙的解决之道,它们会将两层磷脂的尾巴和尾巴相向贴在一起,脑袋都朝外。在这样的情况下,因水分子会极力亲近磷脂分子的亲水脑袋,从而形成巨大的压力,将两层磷脂紧紧挤压到一起,形成一个严严实实的双层“夹心饼干”。
但是,这个过程却远非简单。设想一下,把足量的磷脂分子洒入水中,那就会和洗涤剂的情况一样,形成水面上的一层薄膜,亲水端向水下,拒水端朝上迎向空中。总不能让另一层膜飞到空中降落到它上面将其向下的拒水端与它的向上的拒水端挤压结合到一起吧?
有一种可能,就是有空气搅动的情况下,水下形成很多微小的气泡。气泡被磷脂分子包围,磷脂分子的拒水端朝着气泡内部,亲水端朝外。在周围水的压力下,气泡内的空气被挤出去,于是就形成了两侧亲水端朝外,拒水端挤中间的一张薄膜了。但是这一张平膜怎么又能自己围成一个球形,把水围在中间的呢?
这个物理过程究竟是如何完成的,也许只有上帝知道了。
但正是这样的球体形成了细胞膜的基本形态。这种情况下的细胞膜是依靠水的亲和压力挤压形成的,而不是分子间的化学键形成的。水的亲和压力虽然十分强大,但还是要比化学键弱小不少,因此,可以允许一定的穿凿性。
就这样,既亲水又拒水,既“严”又“疏”两道看起来难以解决的难题,就被磷脂分子破解了。
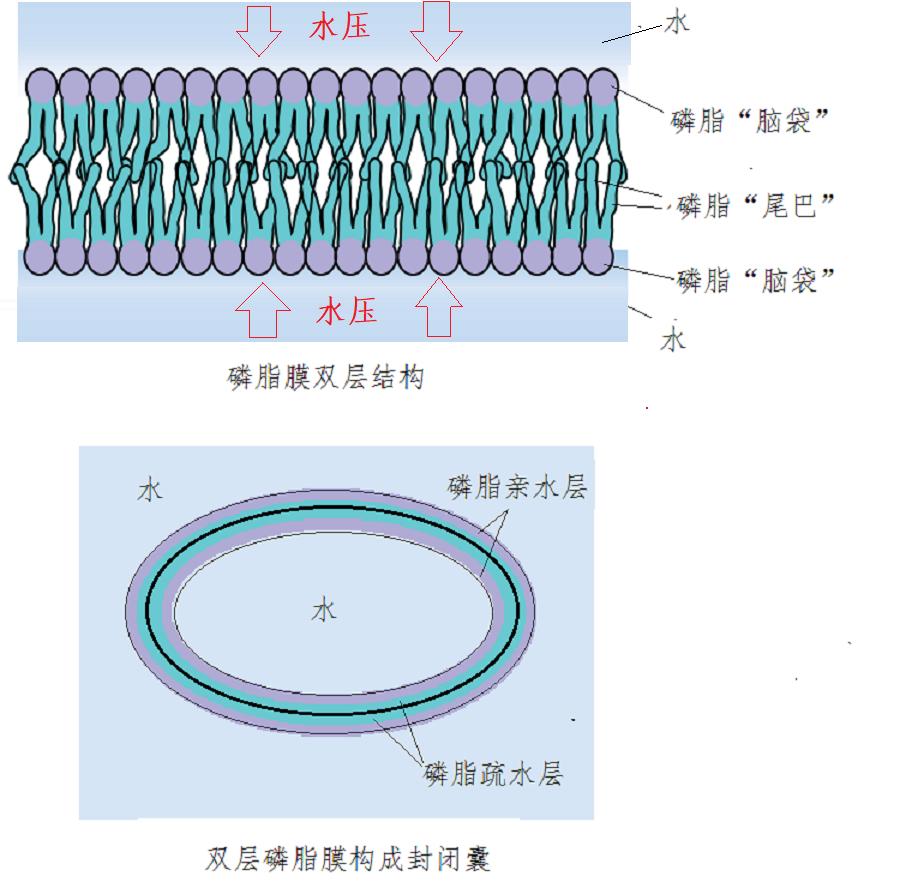
图2磷脂膜形成封闭球囊
二、细胞膜的神奇特征
人们的住房是用砖或石块砌成的,但屋子里的人要维持生命活动还必须有与外界沟通的通道:窗子保持空气流通和光线进入,门让柴米油盐酱醋茶之类的生活物品能搬进来。窗子和门就不能用砖做了,必须用木材或其他材料。
如果说,细胞膜是细胞的围墙,那么,磷脂分子就相当于砌在围墙上的砖,是最基本的了。由磷脂分子筑成的“围墙”必须满足细胞各种各样的活动的需要,要比人们日常生活所需的柴米油盐和其他生活要求要复杂无数倍。
细胞膜厚度约为5纳米。我们日常使用的保鲜膜厚度大约为0.01毫米,已经被认为是非常薄了,然而是细胞膜厚度的2000倍!磷脂分子在细胞膜内外两层的分布并不总是相同的,有的内层多外层少,有的却相反。不同的细胞磷脂的分布也不一样。构成细胞膜的磷脂不止一种,人体细胞膜就至少有四种不同的磷脂分子。
磷脂分子在细胞膜内远不是砌在墙上十分固定,而是会不断移动的。薄膜、纸或墙上绘一些花卉或人物之类的图画,如没人去改动就会始终保持原样不变。如果这些图画自己忽然乱动起来:花瓣自己飞到天上,人脸的眼睛鼻子错位,各处乱跑,甚至正面的图翻到反面,反面的转到正面……此类现象当然都绝不会发生。然而,在细胞膜上却是家常便饭。细胞膜的磷脂分子和附在上面的蛋白质等等都是会“游动”的,犹如不断变幻的马赛克拼图。磷脂分子在膜上的移动速度大约为每秒钟几微米,用不了一秒钟就可以在动物细胞的一端移到另一端。膜上的蛋白质也会移动,速度要较磷脂低一些,但每秒移动的距离也能达到相当于自己“身长”的数百倍马每秒最快跑自己体长的10倍左右(图3)。
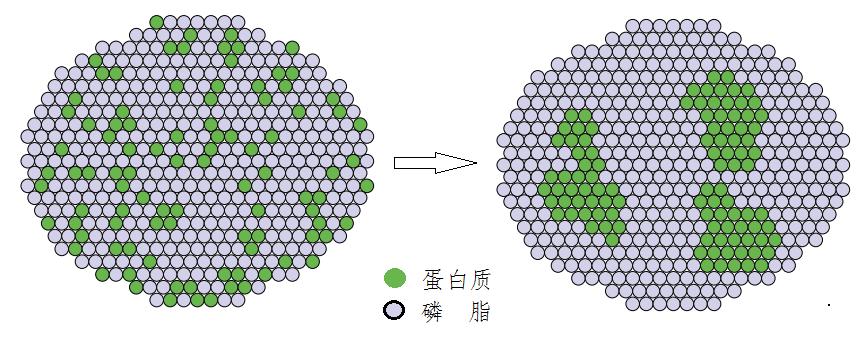
图3蛋白质在细胞膜上移动
磷脂除了沿着细胞膜横向移动,还能从一面翻转到另一面。在人群中挤过去虽然很费劲,但是,通常还能勉强做到,而要在拥挤的人群中翻筋斗,那就匪夷所思了。而磷脂分子也能做到,不过要慢得多,半数磷脂分子翻完筋斗需耗上好几天。而且要靠膜上的翻转酶的帮助才能完成(图4)。
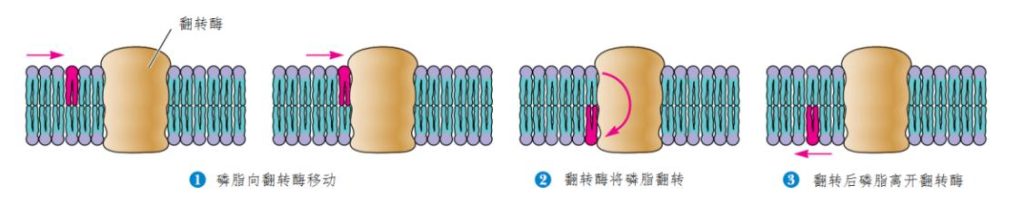
图4 磷脂分子在细胞膜上翻筋斗
磷脂分子的移动不仅单独行动,而且还会集体统一行动。这些流动会有效改变膜的局部功能,如一些插入蛋白质会聚集到一起构成相互作用的网络。因此也很重要的。
细胞膜除了隔离毒物和存储营养,还必须具有传输物质、能量和信息、对外界的反应,以及相互之间联系等等多种功能。我们平常生活用品都通过大门捎带进来,人进出或者客人来访也都通过同一个门,抛弃的物品或垃圾也通过同一大门扔出去。从来都是一门多用的。然而,细胞却往往更多的是一门一用——“大猫钻大洞,小猫钻小洞。”而且要远比“大猫钻大洞,小猫钻小洞。”复杂得多。
细胞的绝大多数对外功能是依靠细胞膜上的蛋白质来完成的。细胞膜上除了磷脂外,还分布各种各样的蛋白质和其他分子(图5)。细胞膜上有很多不同种类的蛋白质,总量基本上能与磷脂相当。这些蛋白质有的粘在细胞膜表面,有的插入膜内,很多不仅插入还两头穿透,甚至往往还不止一次穿过膜。在不同的膜中,蛋白质种类的数量从十多种到超过上百种,发挥主要功能包括,酶、泵、通道、结构组成,抗原和接收功能等等。有的细胞膜上蛋白质总量甚至是磷脂的二倍。
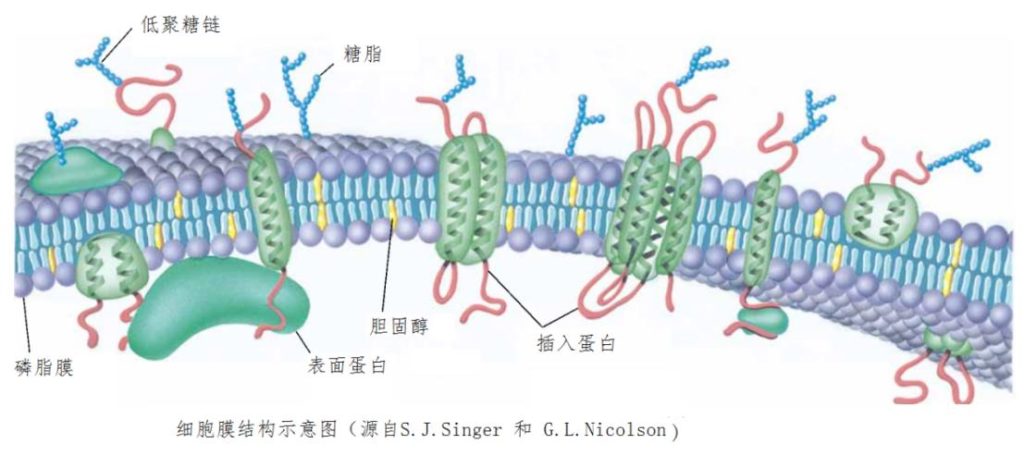
图5细胞膜分布有蛋白质等各种大分子
真核细胞内部还有许多更小的细胞器,也都是由膜包裹起来的。哺乳动物细胞有20多种满足不同功能的膜。
先谈小分子的穿膜输送。磷脂分子两根尾巴中,有一条是弯弯曲曲的,不让磷脂分子彼此挤得太紧密,二氧化碳和氧等中性小分子得以穿透过细胞膜。然而,对于较大体量的分子或者是极性离子就困难了。在自然扩散下,化学物质总是从高浓度处向低浓度处扩散,例如,一颗糖投入到一杯水中,过了些时候,即使没搅动水,整杯水都会甜了。但要用这种方式透过细胞膜扩散,对细胞维持生命来说实在太缓慢(图6)。如果,细胞大量需要的葡萄糖要靠这种扩散方法进入的话,细胞就将饿得无法动弹。
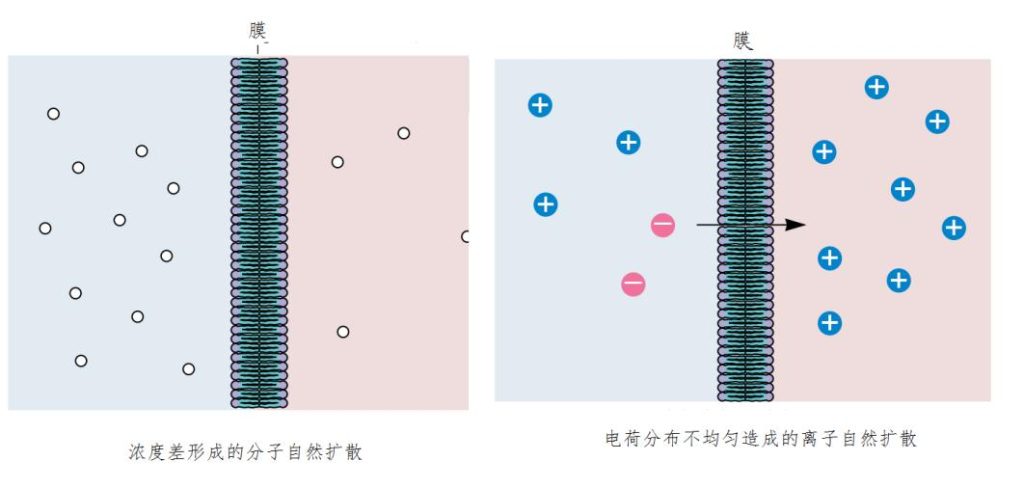
图6 依靠自然扩散速度难以维持生命和活动
然而,细胞膜有多种通道能将输送加速。细胞膜内有各种离子通道,例如:输送钠离子、钾离子和钙离子的阳离子通道,输送氯离子的负离子通道,都是由跨膜蛋白质构成的。这些蛋白质有极复杂的结构,来回穿过膜十多次以上是家常便饭。这些通道还具有很强的针对性和选择性,只允许它选定的离子通过,其他一律拒之门外,而且还都能随时实时调控,根据情况或开或关通道的大门。
一些细胞通道能顺着离子扩散方向强制加速。一种非常有趣的方式是,输送蛋白质会“乒乓式”变换形态,先朝着离子密度高的区域张开,囊入离子后,立即翻转,将离子抛进低浓度区域,就这样不断循环往复(图7)。离子经过通道穿过膜的数量能达到每秒数百万甚至数千万个的惊人程度。尽管如此,匪夷所思的是这种加速却不需要消耗任何能量。
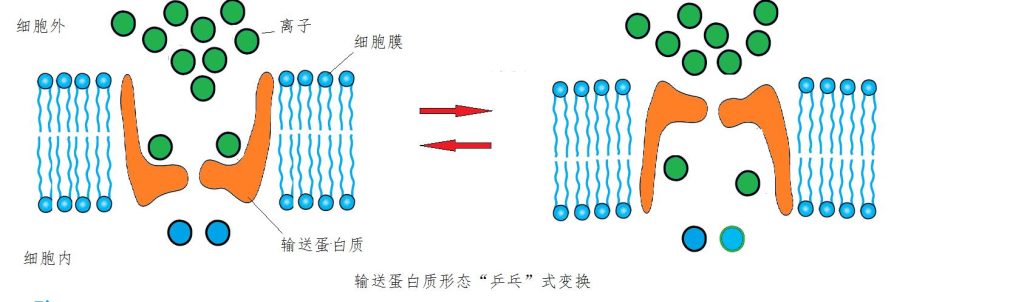
图7 乒乓开闭式细胞通道
细胞膜通道除了能顺势强行加速,还能强行逆势输送。这就需要外加的能量,普遍采用的是一种被称为ATP的高能量分子水解时提供的能量。
例如,钾离子在细胞内能促使许多过程进行,而钠离子则相反,起到抑制作用。所以,动物细胞内钾离子浓度为100mM,细胞外是5~10mM,而钠离子则相反,细胞内浓度为10mM,细胞外是100~140mM。细胞膜上一种名为钠泵的蛋白质通道,源源不断将离子从低浓度处强行反向输送到高浓度处,以维持这种浓度差。钠泵每分解一个ATP,就将2个钾离子送进细胞内,将3个钠离子从细胞内驱赶出去。
钠泵是借助于不断变换自己两种形态,来完成这两种离子的反方向传输的。第一种形态和钠离子比较亲密,结合了3个钠离子。接着就改变成第二种形态,这时候,钠泵“变脸”,“讨厌”了钠离子,反倒对钾离子“亲热”起来,于是,将钠离子释放到细胞外,而将细胞外的2个钾离子拉过来。接着,又再一次“变脸”恢复到第一种状态,对钾离子疏远,从而将钾离子释放到细胞内。就这样不断频繁“变脸”,完成钠离子和钾离子的输送(图8)。
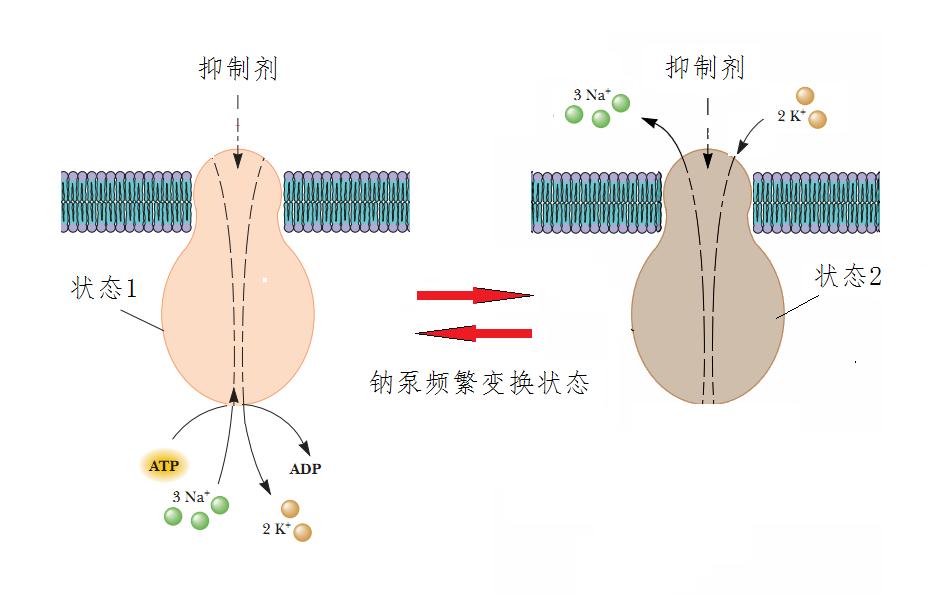
图8 钠泵的运转
钠泵的结构也是非常复杂的,其蛋白质多次穿过细胞膜。图9是把它掰开,平铺开来的示意图。
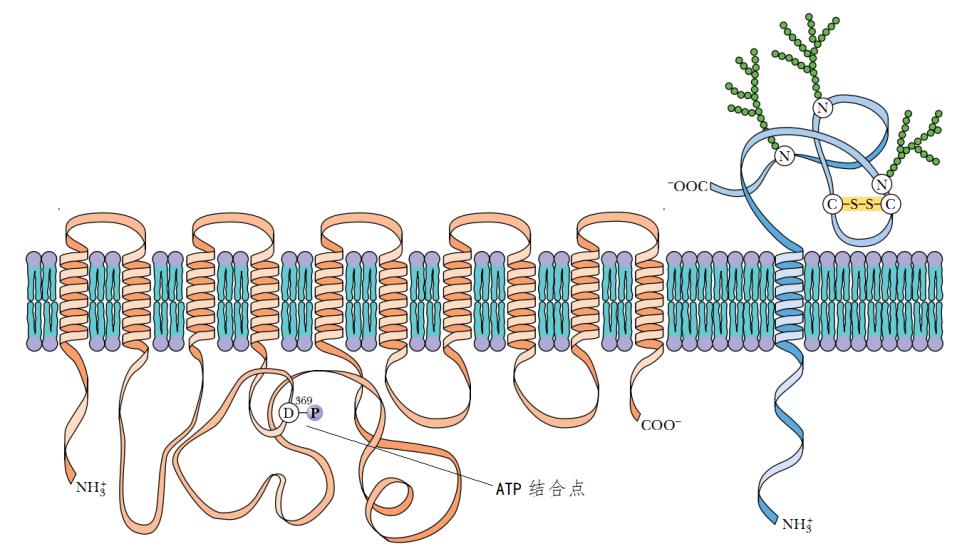
图9钠泵展开图
钠泵“变脸”的速度是非常快的,任何川剧演员变脸的速度都望尘莫及。事实上,动物细胞中ATP能量之20%~40%消耗于此,在神经细胞中甚至高达70%。
除了钠泵还有各种蛋白质构成的输送泵。如:所谓的钙泵能使肌肉纤维周围的浓度瞬间变化上百倍,促使肌肉松弛与收缩之间的转换;以及调整胃液PH值的氢离子和钾离子泵(胃细胞)等等。
除了输送小分子的离子外,还有输送较大分子的通道。例如,提供细胞消耗大量能量的葡萄糖也是通过蛋白质通道输送的。此种通道蛋白质来回穿过细胞膜12次,上面有3个螺旋片段构成输送通道。图10是这种蛋白质展开后的示意图。
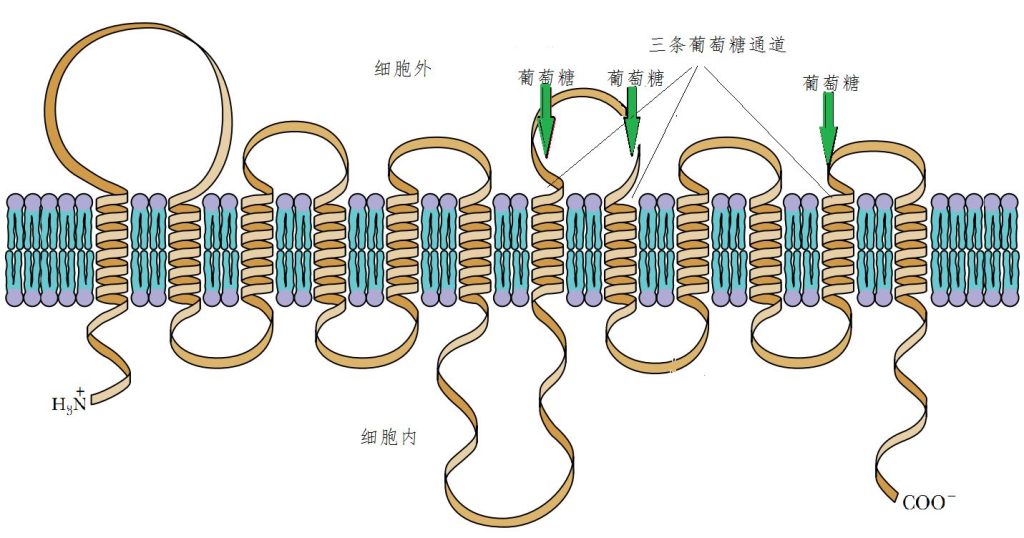
图10葡萄糖细胞膜通道
葡萄糖在向细胞内输送过程中,由于细胞外的钠离子浓度远远高于细胞内的,将葡萄糖“拉”进细胞内的同时钠离子也会顺带进入细胞内。进入细胞内的钠离子会再由钠泵“泵”到细胞外。这种蛋白质通道在红血球细胞上多达5万个,每个每秒钟最高可输送180个葡萄糖分子!也就是说,它们总的可以在一秒钟内向红细胞内输入近1000万个葡萄糖分子1!同样的情况也出现在其他糖类和氨基酸的运输。
家庭除了日常生活外,还得有防盗等自卫功能。细胞膜除了输送功能外,一些细胞膜还能吞噬和吐出分子量较大的分子。例如,高等动物一些巨噬细胞和中性粒细胞等吞噬细胞,能通过吞噬作用消灭感染的细菌、病毒以及损伤的细胞、衰老的红细胞的等等。巨噬细胞的细胞膜每分钟能出现约占3%面积的向内凹陷,每30分钟可遍体都凹陷一遍,每小时能吞下相当于自己体积25%的量。吞下后又重新闭拢。所以,这些细胞虽然没长嘴巴,但遍体又随时随地都还能张嘴(图11)。
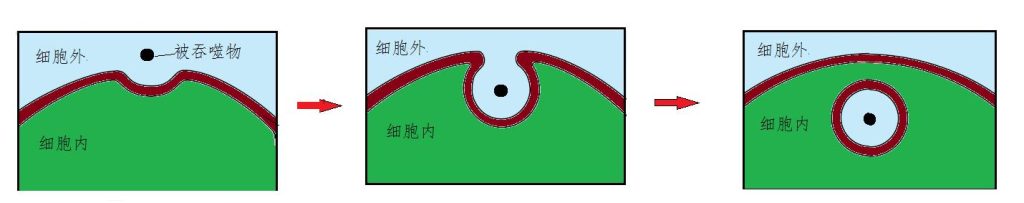
图11 细胞膜吞噬作用
我们生活中常常要与邻居交流接触,互通消息,既有张家长李家短聊天,也有市场信息。而多细胞生物的每个细胞都必须与周围的以及远距离的细胞不断交换信息,以保持密切合作和协调。例如跑步的时候,腿上每个肌肉细胞都必须协同行动,神经纤维不可能连接到每个肌肉细胞上,就靠细胞间互相传递信息。这里有一个明显的例证。将玻璃培养皿中的一个心肌细胞取出来,它的跳动会逐渐失去其原有的同步性。如果再把它放回到培养皿心肌细胞群中,它的跳动又会恢复与细胞群同步。肾细胞生长时会寻找肝细胞,避免与之相接触。细胞一般会自由生长,但一旦与同类细胞接触后生长就会停止,即所谓的接触抑制。癌细胞就是丧失了这种功能,而会疯狂生长。
细胞膜内插入有许多蛋白质组成的六聚体,六聚体中间有一直径为1.8~-2纳米的通道;当两个细胞相互接触后,许多小分子就可以通过这样的通道,从一个细胞转移进入另一个细胞,完成物质或信息的传递。当不需要传递时,两个磷脂膜上的六聚体会相互错开一点,或者,扭转“拧紧”一些,使孔道直径变小。两种情况下,小分子都无法通过,孔道就被关闭了(图12)。
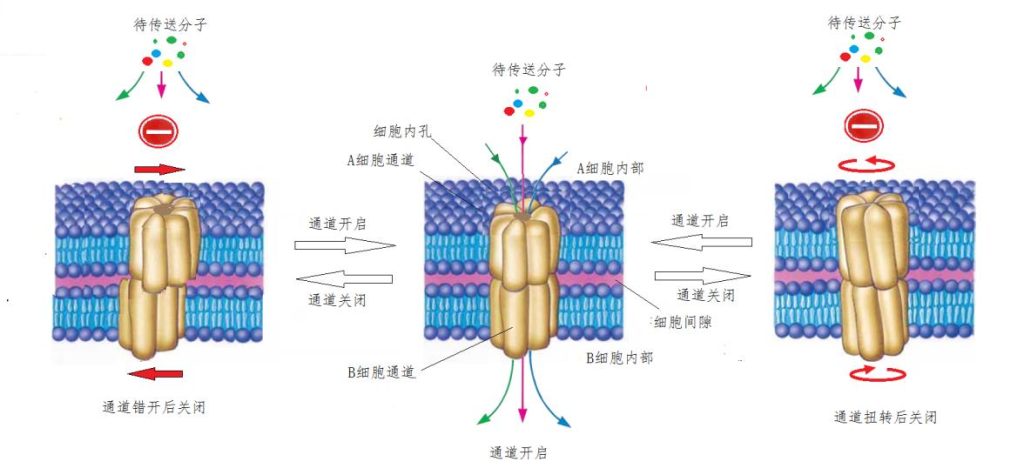
图12 相邻细胞的细胞膜通道的开闭
两个细胞上的孔道必须彼此非常精确对准,传递才可能进行,而且这种彼此瞄准是它们在膜表面上移动非常迅速的情况下完成的。人发的直径为0.05毫米左右,相当于这种孔道直径的2万多倍!没哪个人能夸下海口说,自己能将两根头发丝头对头相互对准,即使是稳坐在椅子上操作。而要将只有头发直径二万分之一的孔道相互快速对准,即使今天最精密的自动机器也未必能做到。
细胞生命的这一切活动所依仗的,都凭借磷脂分子的这种貌似简单却非常巧妙的一个脑袋两根尾巴的基础上。
三、 磷脂的分子结构与生成
任何一个非常高明的设想,如果无法实现,那也只能是放空炮。上面我们看到,细胞膜基本材料的磷脂分子发挥的功能是如此惊人的不可思议。尽管看起来其结构比较简单,但是,要把它们制造出来,难度之大,即使当今诺贝尔化学奖得主都望而却步。现在简单介绍一下。
磷脂的化学结构是由含有三个羟基的甘油与两个脂肪酸和一个磷酸缩合后得到的产物。甘油的第一个碳原子与有16~18个碳原子的饱和脂肪酸结合,第二个碳原子与16~20个不饱和脂肪酸结合,第三个碳原子则与磷酸结合。两根脂肪酸的羧酸与甘油缩合后,尾巴是拒水的碳氢化合物长链。而磷酸部分带有两个带负电荷的氧原子,能与水分子中的带正电荷的氢原子相互吸引,所以能亲水(图13)。
磷酸是无机化合物,在无生命的世界里到处都有,并不稀罕。甘油则不然了,以前从植物中抽取甘油,今天的工业甘油主要以丙烯为原料生产。丙烯是生命体深埋在地下至少数百万年后形成的石油,经高温裂解生产出来的。所以,源头都来自生命体,而不是无机物。真要从无机物直接合成甘油,也不是件简单的事。
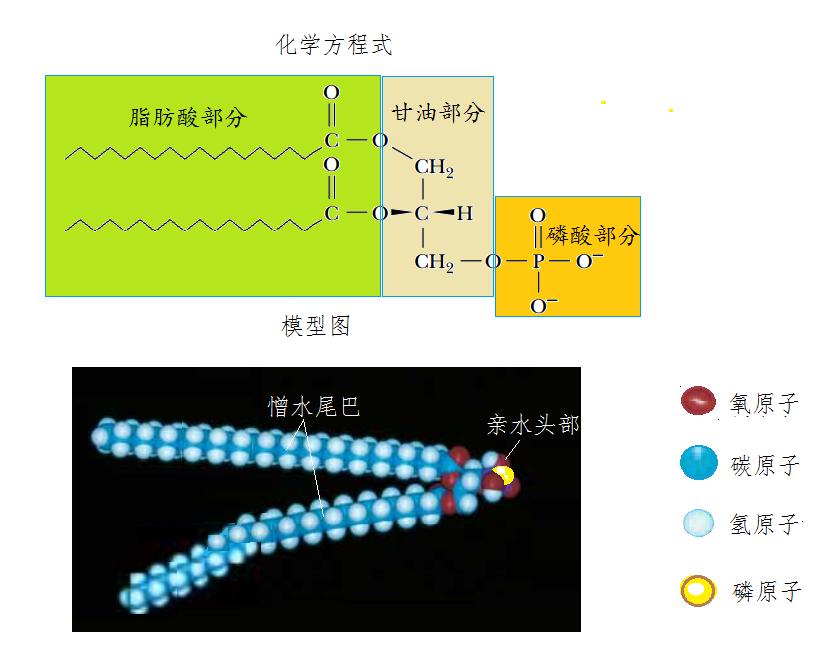
图13 磷脂分子
我们这里仅讨论生命体内磷脂的长链脂肪酸的合成2。单此一项之难度之大,足以让人留下非常深刻的印象。
工业生产中,含有多个碳原子的长链聚合物是用乙烯、丙烯等不饱和分子聚合的。如聚乙烯、聚丙烯等等今天虽然已经是司空见惯,但聚合难度还是相当大的。德国化学家齐格勒发明了用催化剂将乙烯聚合出高分子量的聚乙烯,就是因这方面的杰出贡献,获得了1963年的诺贝尔化学奖。
(即使如此,工业聚乙烯或聚丙烯等的原料乙烯、丙烯还是来源于裂解史前生命形成的石油得到的。)
而且聚合后的产物,还得再氧化才能成为脂肪酸。然而,长链烷烃具有高度的化学稳定性,无论是强碱、强酸、强氧化剂都无法与之发生反应。在高温、光照或催化剂作用下,虽然也能进行一些反应,然而主要还是与卤素进行的卤代反应。
所以,今天的科技水平将简单的小分子一步步合成出脂肪酸也并非办不到,但是要在适合生命生存的温度条件下,高效、高速完成这样的任务,也只能望洋兴叹。
然而,我们许多组织包括肝脏、肾脏、肺和脑等细胞中,都日日夜夜合成脂肪酸。这里,我们仅以分子量相对最小的棕榈酸为例加以说明。棕榈酸有16个碳原子,彼此都用单键相连,其一端的碳原子构成亲水的羧酸,是典型的饱和脂肪酸(图14)。
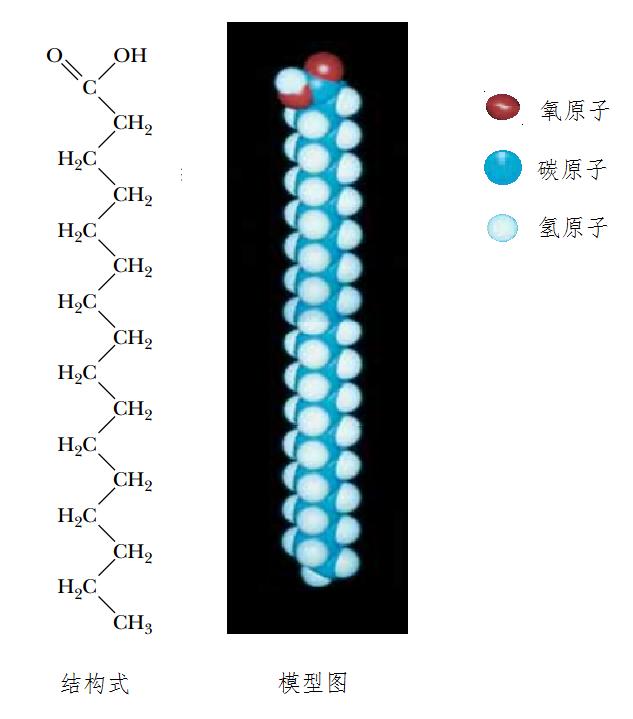
图14棕榈酸分子
要用小分子量的分子去合成含16个碳原子的长链分子,很容易想到的是乙烯、丙烯之类的简单烯烃——只要把它们的双键打开,彼此相连就成了。然而,这些烯烃几乎完全不溶于水,细胞液中没它们踪影。无米之炊做不成。
细胞中是用乙酰为原料的,乙酰同样含有两个碳原子,把8个乙酰的全部16个碳原子都连起来也就可望成功。不过,乙酰不会自己跑去,细胞中是利用辅酶A(CoA)来承担运输任务的(辅酶并不是由蛋白质构成的酶,但是,能对酶的功能能发挥辅助作用,故而称为辅酶。辅酶A具有多种生物功能,今天市场上有很多企业在销售。)辅酶A每次结合一个乙酰,就匆匆赶到聚合脂肪酸合成酶旁,将乙酰基交给合成酶后就离开。
事情才开始。因为,每个乙酰都带有一个氧原子,而棕榈酸的15个碳原子上一个氧原子都没有,所以得将那些多余的氧原子都除掉,这可需要多道步骤。细菌和植物是用多个单独的酶,一个接一个相继执行不同的任务,最后才完成。而哺乳动物、鸟类等的脂肪酸合成是在同一个合成酶内完成的,效率当然要高得多。这种合成酶是由两个完全相同的单元构成的二聚体。每个单元包括7个酶活性区和一个载乙酰的蛋白ACP(图15)。
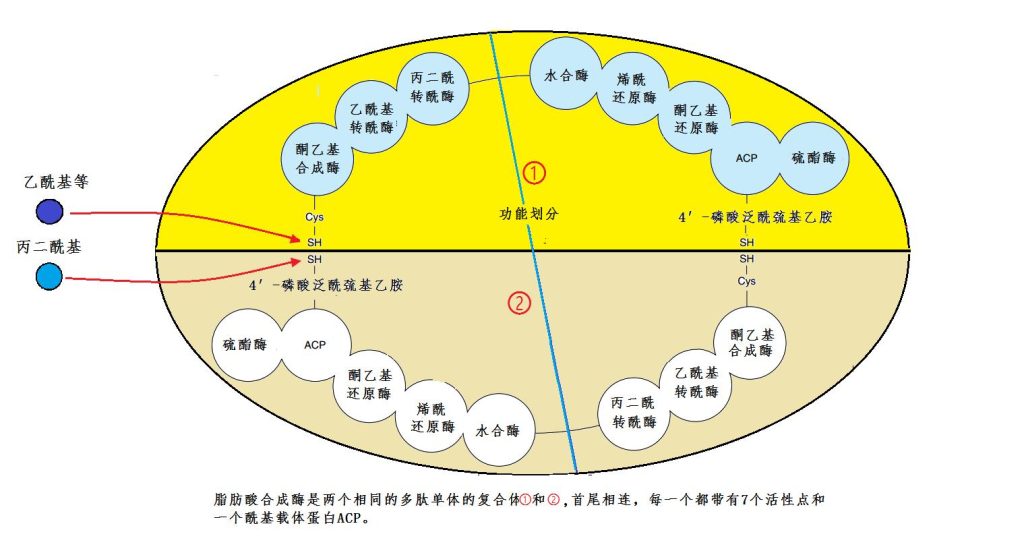
图15 棕榈酸合成酶
合成酶单元2一次次从辅酶A获得乙酰基,然后单元1将附着在自己身上的碳链,加到单元2的这个乙酰基上,于是,单元2的碳链就比单元1的多出两个碳原子。
由于乙酰基上原来是有氧原子的,必须去除。这时候另一种辅酶NADPH(还原型辅酶Ⅱ)和H+登场,提供氢原子与氧原子结合,形成水分子后脱离,然后再进一步将不饱和双键加氢饱和。
如果碳链上的碳原子还不足16个,单元1和单元2就转换角色,继续从辅酶A接受乙酰基团,直到碳链上有了16个碳原子后,才最后水解成棕榈酸,离开合成酶。
这样的过程反复进行7次,每次6步的反应,加上开头与结束的反应,共44步!除了合成酶,辅酶A参与了8次,辅酶NADPH和H+各参与了14次!(图16)
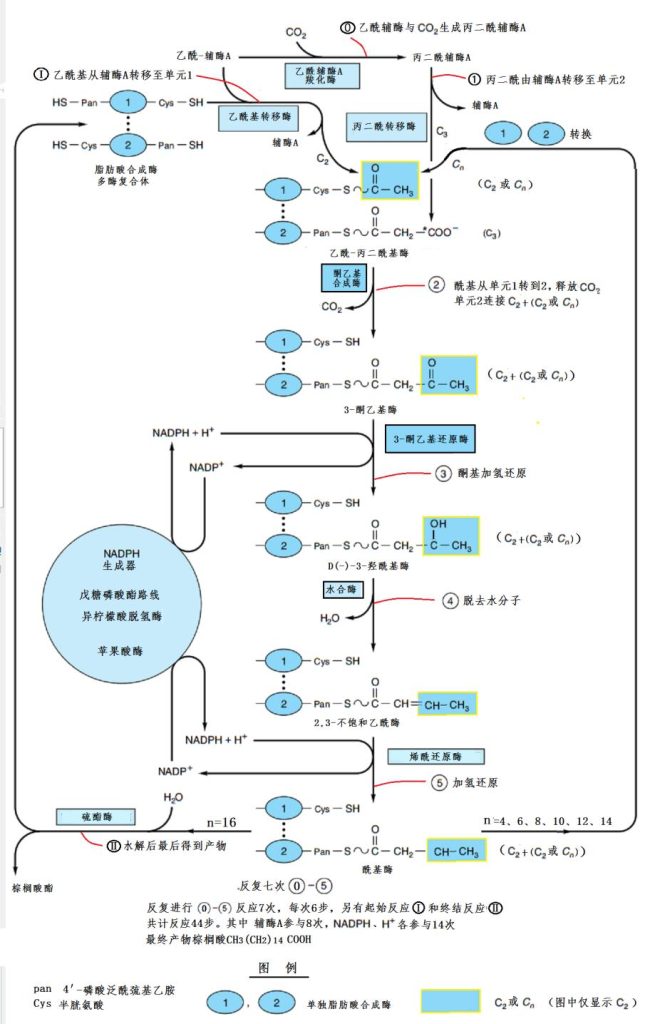
图16 脂肪酸的生物合成路线
(图15和图16均译自参考文献2)
对于化学家来说,即使知道如此合成路线,按照现有的技术水平依然面临一道道巨大难关,如:产物必须只有一种分子量——例如,或者全都是16个碳原子,或者全都是18个碳原子等等,然而人工聚合都有一定分子量分布,要做到统统只有一种分子量又谈何容易;又如,在脱氧过程而生成水的反应,在水环境中又将遇到化学平衡的水魔咒难以逾越的障碍,等等。
即使这些难关能闯过,收率问题也是绕不过去的。今天实验室里有机化学反应的收率能达到70%-80%就很满意了,如果能达到90%就是非常高的了。这里,即使每次收率都能达到90%,44步反应的总收率就只有0.944,连1%都不到了。这还仅仅是磷脂上的一根饱和脂肪酸棕榈酸,另外还有一根碳原子更多的不饱和脂肪酸也必须聚合。棕榈酸的碳原子是16个,磷脂两根脂肪酸的碳原子可高达38个,至少必须进行类似的100多步反应。它们的原料乙酰也必须经过生命体内一系列反应才能得到,两根脂肪酸和磷酸还必须与甘油进行酯化反应,才能形成磷脂。而甘油也不会凭空降临,也必须合成。所以,每合成一个磷脂至少要超过120多步化学反应!即使每次反应的收率都有90%,120步反应的总收率就只有0.9120 约等于0.000003229……,好不容易拿到一个合格的磷脂却将淹没在30多万个杂物中,怎么让它们排除万难聚拢到一起,往下还怎么玩?
而脂肪酸和磷酸与甘油的酯化缩合还有两道难关:首先是长链脂肪酸往往都是不溶于水的,如何去与水中的甘油反应?其次是这样的反应将释放出水,这又面临水魔咒的化学平衡的问题。这里,或许有人会问,那么生命是如何战胜水魔咒的呢?对这个问题,我们在以后章节中再展开来谈。
而生命却极高效地完成了看来几乎不可能完成的任务。以大肠杆菌为例,每个大肠杆菌的细胞膜有数千万个磷脂分子,细菌是用多个单独的合成酶,一个接一个相继执行不同的任务,最后才完成。大肠杆菌平均每20多分钟繁殖一次, 20多分钟内合成了数千万个磷脂分子,所以,大肠杆菌每分钟内进行数亿次的化学反应!真核生物细胞的尺度是大肠杆菌的10~100倍,细胞膜的磷脂分子数量动辄要达到数千亿之多。哺乳动物的受精卵一昼夜左右就分裂成两个,数千亿的磷脂分子必须在短短的一昼夜中统统制造出来。
最后,乙酰、乙酰辅酶A, NADPH以及脂肪酸合成酶都是生命产生的产物。生命细胞必须有膜包裹,而膜和其原料又必须在细胞内制造。我们看来陷入了一个循环论证困境——一个典型的先有鸡还是先有蛋的问题。以后,我们将看到生命现象无数次这样的情况。
我们对不太了解的事情常会说“略知皮毛”,细胞膜不过是细胞的“皮毛”而已,而磷脂则更是“皮毛”上的微小颗粒。而对这样的“皮毛颗粒”今天的高科技还依然望尘莫及。
看了这些,还能相信这一切都是偶然产生的吗?但是,如果是智慧的产物,那么,那是何种样的伟大智慧啊!
以后还要介绍更多复杂巧妙得多的生命现象。
参考文献:
1.顾德兴等,《普通生物学》,高等教育出版社,2000年,p39
2.《Harper;s Illustrated Biochemistry》By the McGraw-Hill Company Inc.2003year ,P173-179,415-430


